
Dineina: struttura e classi funzionali
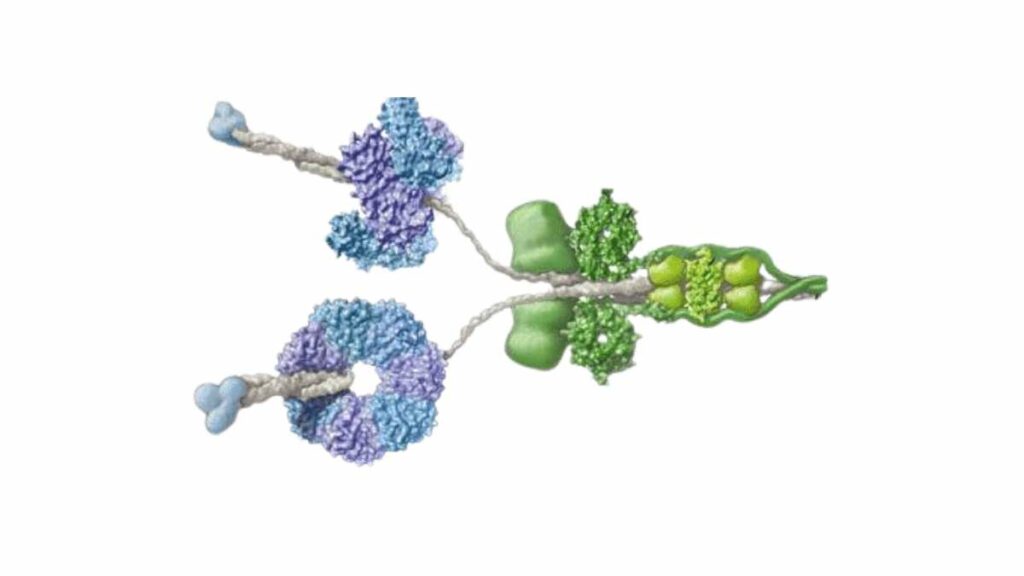
Non è semplice studiare la dineina a causa della sua struttura articolata che riunisce diverse subunità. Si tratta di una proteina dalle proprietà particolari che in molti libri di scuola è indicata come il “motore proteico” della cellula. Con questo nome però non si indica una sola molecola ma addirittura due classi di polipeptidi, divise in base alle funzioni e localizzazione che hanno.
Per muoversi questa proteina ha bisogno di interagire con i microtubuli del citoscheletro cellulare. Si può dire che si tratta dei binari che deve percorrere, e la direzione che segue è sempre la stessa. I microtubuli hanno infatti un’estremità positiva e una negativa, di cui la seconda è quella che dà il verso in cui orientare il movimento a questa proteina.
La struttura della dineina
Per funzionare questa classi di proteina necessita di ATP, di cui può legare quattro molecole in tutto. Il cuore della sua struttura è composto in tutto da sei domini chiamati AAA+ (ATPasi Associate ad Attività) numerati da 1 a 6. In più sono presenti un linker che tiene coesa la proteina e una coda (tail) che protrude dal core e può legare la molecola da trasportare lungo i microtubuli.
Il movimento origina dall’idrolisi dell’ATP che avviene nel dominio AAA1, l’unico che può catalizzare questa reazione. Gli altri 3 ATP legati alla dineina infatti non vanno incontro a questa reazione e sembra abbiano solo una funzione regolatoria. L’idrolisi produce un movimento a scatto che si propaga dal dominio AAA1 fino all’AAA4. Questo dominio presenta un lungo stelo, che ruotando sposta in avanti la proteina.
Lo stelo è infatti la parte strutturale che collega questo motore proteico al suo “binario”. Una volta che AAA1 lega una nuova molecola di ATP il movimento si ripete. Secondo quanto si è ricavato dalle analisi al microscopio elettronico lo stelo della proteina si lega al microtubulo tra due dimeri consecutivi. La tubulina che li compone infatti è formata da dimeri ripetuti delle subunità α e β.
Ogni movimento o “passo” porta lo stelo ad avanzare di circa due dimeri di tubulina, una distanza pari a 16 nm. La molecola da portare all’estremità del microtubulo però essendo legata in un altro punto si muove della metà, quindi 8 nm.
Le due classi funzionali
Come precisato con il nome dineina si indicano più proteine, che si possono raggruppare in due classi funzionali distinte. Quella che abbiano trattato al paragrafo precedente è la classe citosolica, in quanto si tratta di proteine che operano nel citoplasma delle cellule. La loro funzione principale è il trasporto intracellulare di più tipologie di “carichi”.
All’interno della cellula sappiamo che avviene continuamente movimento di vescicole contenenti enzimi, proteine o altre molecole. Si tratta di piccole strutture delimitate da uno strato fosfolipidico (semplice o doppio) per proteggere il contenuto dal pH del citosol. Possono derivare dalla gemmazione del Golgi o provenire dall’ambiente esterno alla cellula.
Oltre alle vescicole la dineina può trasportare anche organelli interi, come ad esempio i mitocondri. La seconda classe funzionale invece è quella che raggruppa le cosiddette proteine dell’assonema. Con questo termine si indica la struttura interna delle ciglia e dei flagelli, protrusioni presenti in alcune tipologie di cellule che possono muoversi.
Ciglia e flagelli differiscono prima di tutto per la lunghezza dato che i secondi sono più lunghi, ma anche per numero. Una cellula ciliate presenta diverse strutture di questo tipo mentre una flagellata di solito ha una sola protrusione. All’interno hanno una complessa struttura di microtubuli oltre a proteine per il trasporto, tra cui le chinesine.
La struttura dell’assonema
In ciascuna delle coppie di microtubuli dell’assonema ce n’è sempre uno più spesso (A) composto da tredici microfilamenti, come vale per quelli del citoscheletro. Quello adiacente invece ne presenta solo dieci nella sua struttura ed è leggermente più sottile(B). L’intero assonema è circondato da una membrana plasmatica che è in continuità con quella del corpo cellulare.
L’estremità positiva delle coppie di microtubuli interne al flagello è rivolta verso l’interno ovvero sul punto più vicino alla cellula. In questo modo la dineina, muovendosi verso l’estremità negativa, si allontana dal corpo cellulare verso la punta del flagello. Per spostarsi la proteina si muove lungo il lato del microtubulo che guarda la membrana plasmatica, non quello interno che è rivolto verso la coppia centrale.
Non la si trova su entrambi i microtubuli della coppia ma solo su quello completo. Dalla coppia centrale dell’assonema partono delle strutture dette proiezioni che la collegano a tutte le doppiette che la circondano. Dove avviene l’attacco del flagello alla cellula si trova il corpo basale, una struttura dove i microtubuli sono presenti a gruppi di tre (ma per ogni tripletta uno è più corto degli altri).

